
Где в митохондриях происходит синтез атф в крестах
Обновлено: 06.07.2024
Строение и функционирование F1F0-АТФ-синтазы
Основная масса энергии в клетках эукариот и почти всех прокариот запасается в процессе окислительного фосфорилирования в форме АТФ. Этот многоступенчатый процесс происходит в мембранах.
В организме человека весом около 70 кг за 75 лет при сидячем образе жизни нарабатывается 2 млн кг АТФ. В клетках эукариот синтез АТФ, который является одним из наиболее распространенных ферментативных биологических процессов, происходит в специальных органеллах, называемых митохондриями.
Образование АТФ в клетке катализируется ферментом F1F0-АГФ-синтазой, который представляет собой необычный молекулярный мотор, использующий энергию электрохимического градиента протонов (трансмембранный протонный градиент) для синтеза АТФ. Электрохимический градиент протонов создается комплексами, переносящими электроны в ходе окислительного фосфорилирования.
Он определяется двумя показателями: мембранным потенциалом и разностью концентраций протонов по сторонам мембраны митохондрий.
Общая структура F1F0-АТФ-синтазы во всех клетках одинакова, хотя состав субъединиц может различаться. Простейшая форма F1F0-АТФ синтазы цитоплазматической мембраны бактериальной клетки состоит из восьми типов субъединиц и обладает молекулярной массой 530 кДа.
Состав ее может быть выражен стехиометрической формулой а3 b3 γ δ ε a b2 с10-14. В митохондриях фермент содержит 7-9 дополнительных регуляторных субъединиц, которые немного увеличивают его молекулярную массу. АТФ-синтаза состоит из двух доменов:
• связанный с мембраной домен F0 (ab2c10-14 у бактерий), который участвует в транслокации протонов в направлении электрохимического градиента, и
• глобулярный домен F1(а3 β3 γ δ ε у бактерий), содержащий каталитические сайты, ответственные за синтез АТФ. (Изолированный F1 домен обладает способностью к гидролизу АТФ.)
Субъединицы с домена F0 образуют кольцо, которое взаимодействует с субъединицей а. Субъединица у формирует центральный выступ ротора, который связан с основанием кольца с и входит в каталитический домен F1. Этот домен состоит из трех а и трех b субъединиц, которые расположены попеременно и образуют структуру гексагонального цилиндра вокруг субъединицы у.
Выступающая ножка статора состоит из субъединиц b2δ, причем субъединица δ связана с доменом F1, а b2 с доменом F0 митохондриальной мембраны и с субъединицей а.
Каким образом F1F0-АТФ-синтаза использует энергию трансмембранного протонового градиента для синтеза АТФ? Предложенная модель постулирует существование следующих основных этапов:
• субъединицы а и с контролируют транспорт протонов таким образом, что с-кольцо вращается по отношению к субъединице а. Иными словами, энергия электрохимического градиента протонов расходуется на механическое вращение субъединиц с;
• у-субъединица центрального выступа вращается вместе с с-кольцом, при этом трансмембранная движущая сила протонов передается на расстояние 100 А домен F1; и
• механическая энергия вращения используется для высвобождения АТФ, синтез которого катализируется b-субъединицами домена F1.
Поэтому вращение с-кольца и центральной у-субъединицы относительно субдомена а3 b3 играет существенную роль в сопряжении трансмембранной движущей силы протонов с образованием и выходом АТФ. Поскольку каждая с-субъединица несет один протон, то при полном обороте с-кольца (в зависимости от типа АТФазы) транспортируются 10-14 протонов, и на одну молекулу образующегося АТФ транспортируются около четырех протонов.
F1F0-АТФ-синтаза превращает электрохимическую энергию в механическую и снова в химическую, почти со 100%-ной эффективностью. Синтез АТФ может проходить с максимальной скоростью порядка 100 с-1, и уровень АТФ в клетке может достигать миллимолярных концентраций.
У некоторых бактерий F1F0-АТФ-синтаза работает в обратном направлении, и энергия, высвобождающаяся за счет гидролиза АТФ, расходуется на выход протонов из клетки и на создание их градиента по сторонам цитоплазматической мембраны. Затем энергия протонного градиента используется для транспорта в клетку метаболитов, например лактозы.
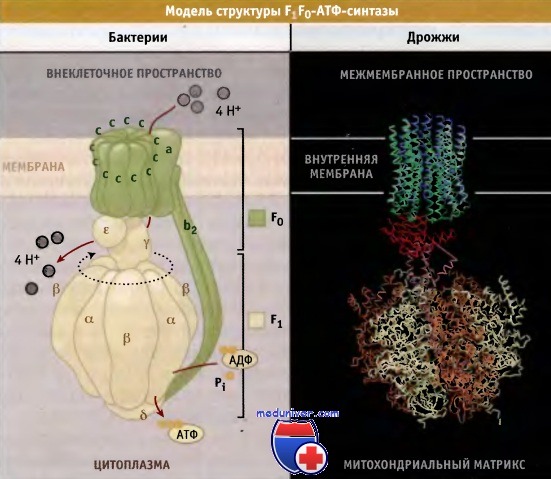
Схематическое изображение комплекса F1F0-АТФ-синтазы Е. coli и часть кристаллической структуры дрожжевого фермента.
Изображение кристаллической структуры дрожжевого фермента построено на основании Protein Data Bank file 1Q01.
Показано предполагаемое положение мембраны.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Механизмы образования АТФ в митохондриях
Все живые клетки обладают способностью превращать энергию окружающей среды в общий интермедиат — АТФ.
Клетка получает энергию за счет пищи, которую она усваивает из окружающей среды. Затем эта энергия должна быть превращена в форму, которую можно распределять по клетке. Общее решение этого вопроса (не только для митохондрий, но и для прокариотических клеток, также превращающих энергию) состоит в запасании энергии в форме универсального вещества, которое можно использовать в любой момент и в любом месте клетки.
Различные типы клеток отличаются деталями энергетической системы, однако общим свойством, характерным для всех клеток, является их способность превращать энергию окружающей среды в АТФ, который представляет собой общую молекулу, при необходимости обеспечивающую энергией отдельные химические реакции.
АТФ образуется двумя путями: в цитозоле и в митохондриях. Первый путь реализуется в цитозоле эукариотических клеток и у бактерий. При этом в результате гликолиза происходит распад глюкозы с образованием пирувата и образуются две молекулы АТФ. Эта реакция может происходить в анаэробных условиях (в отсутствие кислорода).
Второй путь является основным источником создания энергетических запасов и осуществляется в митохондриях эукариотических клеток. Процесс, при котором в митохондриях образуется АТФ, называется окислительным фосфорилированием и осуществляется с участием электрон-транспортной цепи. Пируват, высвобождающийся при гликолизе, поступает в матрикс (люмен) митохондрии, где он распадается и, реагируя с коэнзимом А, образует ацетил-КоА.
После этого, при участии цикла лимонной кислоты с освобождением атомов водорода, ацетильный остаток ацетил-КоА распадается, образуя двуокись углерода. Восстановление переносчика НАД+ в НАДН происходит при участии атомов водорода, и затем при окислении НАДН высвобождаются протон и электрон. Из матрикса ионы водорода (протоны) переносятся через мембрану в межмембранное пространство, а электроны перемещаются вдоль мембраны с участием нескольких транспортных белков. В результате по обе стороны мембраны создается градиент концентрации протонов.
Под действием этого градиента протоны переносятся через мембрану в обратном направлении. Это осуществляется при участии большого белкового комплекса АТФ-синтазы, которая и образует АТФ из АДФ и неорганического фосфата. Этот процесс носит название хемиосмос.
Митохондрии часто называют энергетическими центрами (эукариотических) клеток; они снабжают клетку энергией, необходимой для осуществления метаболических процессов и структурных изменений. Точнее говоря, они превращают энергию, поступающую из внешней среды в формы, которые могут быть использованы клеткой. Постоянство структуры и функции митохондрий у всех клеток эукариот убеждает в том, что эндосимбиоз, в результате которого они возникли, должен был произойти на самом начальном этапе эволюции эукариот.
АТФ образуется при хемиосмосе с участием системы протонного транспорта.
Видео урок цикл Кребса
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Строение и функционирование F1F0-АТФ-синтазы
Основная масса энергии в клетках эукариот и почти всех прокариот запасается в процессе окислительного фосфорилирования в форме АТФ. Этот многоступенчатый процесс происходит в мембранах.
В организме человека весом около 70 кг за 75 лет при сидячем образе жизни нарабатывается 2 млн кг АТФ. В клетках эукариот синтез АТФ, который является одним из наиболее распространенных ферментативных биологических процессов, происходит в специальных органеллах, называемых митохондриями.
Образование АТФ в клетке катализируется ферментом F1F0-АГФ-синтазой, который представляет собой необычный молекулярный мотор, использующий энергию электрохимического градиента протонов (трансмембранный протонный градиент) для синтеза АТФ. Электрохимический градиент протонов создается комплексами, переносящими электроны в ходе окислительного фосфорилирования.
Он определяется двумя показателями: мембранным потенциалом и разностью концентраций протонов по сторонам мембраны митохондрий.
Общая структура F1F0-АТФ-синтазы во всех клетках одинакова, хотя состав субъединиц может различаться. Простейшая форма F1F0-АТФ синтазы цитоплазматической мембраны бактериальной клетки состоит из восьми типов субъединиц и обладает молекулярной массой 530 кДа.
Состав ее может быть выражен стехиометрической формулой а3 b3 γ δ ε a b2 с10-14. В митохондриях фермент содержит 7-9 дополнительных регуляторных субъединиц, которые немного увеличивают его молекулярную массу. АТФ-синтаза состоит из двух доменов:
• связанный с мембраной домен F0 (ab2c10-14 у бактерий), который участвует в транслокации протонов в направлении электрохимического градиента, и
• глобулярный домен F1(а3 β3 γ δ ε у бактерий), содержащий каталитические сайты, ответственные за синтез АТФ. (Изолированный F1 домен обладает способностью к гидролизу АТФ.)
Субъединицы с домена F0 образуют кольцо, которое взаимодействует с субъединицей а. Субъединица у формирует центральный выступ ротора, который связан с основанием кольца с и входит в каталитический домен F1. Этот домен состоит из трех а и трех b субъединиц, которые расположены попеременно и образуют структуру гексагонального цилиндра вокруг субъединицы у.
Выступающая ножка статора состоит из субъединиц b2δ, причем субъединица δ связана с доменом F1, а b2 с доменом F0 митохондриальной мембраны и с субъединицей а.
Каким образом F1F0-АТФ-синтаза использует энергию трансмембранного протонового градиента для синтеза АТФ? Предложенная модель постулирует существование следующих основных этапов:
• субъединицы а и с контролируют транспорт протонов таким образом, что с-кольцо вращается по отношению к субъединице а. Иными словами, энергия электрохимического градиента протонов расходуется на механическое вращение субъединиц с;
• у-субъединица центрального выступа вращается вместе с с-кольцом, при этом трансмембранная движущая сила протонов передается на расстояние 100 А домен F1; и
• механическая энергия вращения используется для высвобождения АТФ, синтез которого катализируется b-субъединицами домена F1.
Поэтому вращение с-кольца и центральной у-субъединицы относительно субдомена а3 b3 играет существенную роль в сопряжении трансмембранной движущей силы протонов с образованием и выходом АТФ. Поскольку каждая с-субъединица несет один протон, то при полном обороте с-кольца (в зависимости от типа АТФазы) транспортируются 10-14 протонов, и на одну молекулу образующегося АТФ транспортируются около четырех протонов.
F1F0-АТФ-синтаза превращает электрохимическую энергию в механическую и снова в химическую, почти со 100%-ной эффективностью. Синтез АТФ может проходить с максимальной скоростью порядка 100 с-1, и уровень АТФ в клетке может достигать миллимолярных концентраций.
У некоторых бактерий F1F0-АТФ-синтаза работает в обратном направлении, и энергия, высвобождающаяся за счет гидролиза АТФ, расходуется на выход протонов из клетки и на создание их градиента по сторонам цитоплазматической мембраны. Затем энергия протонного градиента используется для транспорта в клетку метаболитов, например лактозы.
Схематическое изображение комплекса F1F0-АТФ-синтазы Е. coli и часть кристаллической структуры дрожжевого фермента.
Изображение кристаллической структуры дрожжевого фермента построено на основании Protein Data Bank file 1Q01.
Показано предполагаемое положение мембраны.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Анаэробное расщепление глюкозы в целом энергетически невыгодно. В связи с этим многие прокариоты и большинство эукариот приспособились использовать появившейся в атмосфере Земли кислород для более полного расщепления (окисления) органических веществ.
Для этого ферментные системы катализируют целую цепь последовательных превращений органических веществ (субстратов) до их полного окисления до углекислого газа (С02) и воды (Н20). Окислительно-восстановительные реакции клеточного дыхания по своей сути представляют собой перенос электронов от окисляемого вещества (донора — органического вещества) к окислителю (акцептору — переносчику водорода). В результате формируется мембранный градиент ионов водорода — ДН+. При достижении определенного трансмембранного электрохимического потенциала (примерно 200 мВ), ионы переносятся через мембрану с помощью фермента АТФ-синтазы и осуществляется синтез АТФ.
Этот процесс называют мембранным окислительным фосфорилированием или клеточным дыханием.
Полное уравнение образования АТФ при дыхании выглядит следующим образом:

По сравнению с гликолизом при кислородном окислении образуется в 18 раз больше АТФ (при гликолизе — 2 молекулы АТФ). Таким образом — полное расщепление одной молекулы глюкозы дает энергию для синтеза 38 молекул АТФ (2600 кДж). КПД всего этого процесса составляет 40 %, а 60 % рассеивается в виде тепла.
У прокариот клеточное дыхание проходит на наружной плазматической мембране, а у эукариот — на мембранах специальных клеточных органоидов — митохондрий.
Митохондрии. Из курса общей биологии вы помните, что митохондрии — характерный органоиды всех эукариотических клеток (растений, животных, грибов) обладающие универсальным строением (рис. 4.14).

Рис. 4.14. Строение митохондрии и основные типы митохондрий:
- 1 — наружная митохондриальная мембрана; 2 — внутренняя митохондриальная мембрана; 3 — матрикс; 4 — митохондриальные рибосомы; 5 — мит-ДНК;
- 6 — включения (гликоген, соли тяжелых металлов, пигменты); 7 — АТФ- синтетазный комплекс; 8 — одиночные митохондрии; 9 — нитчатые митохондрии; 10 — цепочечные митохондрии; 11 — ретикулярные (сетчатые)
Митохондрии имеют двойную мембрану. Наружная мембрана гладкая, внутренняя образует систему впячиваний — крист. Кристы могут иметь разную форму (пластинчатую, трубчатую) и положение (поперечное, продольное). Между мембранами находится межмембранное пространство шириной около 10—20 нм.
Наружная и внутренняя мембраны митохондрий довольно сильно различаются по своему составу и свойствам. Так, наружная мембрана обладает большой проницаемостью для крупных молекул (например; белков, молекул АДФ и АТФ), тогда как внутренняя мембрана пропускает только мелкие молекулы и ионы, а для транспорта крупных молекул имеет систему специальных переносчиков. Основная задача внешней мембраны — избирательно отделить митохондрию от цитоплазмы, а задача внутренней мембраны — обеспечить работу дыхательной цепи для синтеза АТФ. Центральную часть митохондрии занимает митохондриальный матрикс, в котором располагается генетический аппарат митохондрии в виде нескольких кольцевых молекул ДНК; рибосомы (похожие на рибосомы прокариот); ферменты цикла трикарбоновых кислот (цикла Кребса); ферменты для собственного митохондриального белкового синтеза и синтеза РНК и ДНК; гранулы гликогена или крахмала; различные включения.
Митохондрии обладают собственным геномом и аппаратом белкового синтеза, но их автономия неполная. Гены митохондриальной ДНК кодируют примерно 30 % белков, входящих в состав митохондрии. Большая часть структурных и регуляторных белков, ферментов дыхательной цепи, белков рибосом кодируются в ядерном геноме и синтезируются в цитоплазме, транспортируясь затем в митохондрию.
Тем не менее, обладание собственным геномом обеспечивает митохондриям уникальное свойство — способность к делению. Перед циклом деления происходит репликация ДНК, а затем митохондрии делятся пополам или отделяется фрагмент, содержащий ДНК (так называемое почкование митохондрий).
В эукариотических клетках встречается несколько разновидностей митохондрий. Во многих клетках (одноклеточные организмы, клетки хрящевой ткани, клетки печени, секреторные клетки поджелудочной железы и др.) присутствуют многочисленные мелкие отдельные митохондрии, рассеянные по цитоплазме.
В других клетках (нейронах, лимфоцитах, клетках соединительной ткани, у одноклеточных водорослей) митохондрий немного, и они представляют собой систему одиночных сильноразветвленных органоидов. В поперечно-полосатых мышцах позвоночных и беспозвоночных животных, требующих большое количество энергии, митохондрии образуют целую сеть огромных разветвленных митохондрий. В сердечных мышцах отдельные мелкие митохондрии соединены между собой системой специальных контактов и также образуют разветвленную внутриклеточную митохондриальную сеть. Профессор Московского государственного университета Ю. С. Ченцов, подробно исследовавший различные типы митохондрий, предположил и экспериментально доказал, что одиночные разветвленные митохондрии и митохондриальные сети предназначены для бесперебойного и синхронного синтеза АТФ и доставки его в любую удаленную часть клетки (например, вдоль миофибриллы соматического мышечного волокна или цепочки мышечных клеток сердца, или длинного отростка нервной клетки).
Синтез АТФ. Синтез АТФ идет на внутренней мембране митохондрий, где сосредоточен комплекс ферментов дыхательной цепи и ферменты синтеза АТФ. Основные этапы этого процесса согласно хемио- осмотической теории П. Митчелла (лауреата Нобелевской премии) можно представить себе следующим образом (рис. 4.15):
1) после первого этапа энергетического обмена — гликолиза, проходящего в цитоплазме, образуются две молекулы пировиноградной кислоты (С3Н4О3) и две молекулы НАД • Н2 (еще один восстановленный переносчик атомов водорода). Пировиноградная кислота поступает в матрикс митохондрий где окисляется, здесь, При этом образуется молекула активированной уксусной кислоты (ацетил-коэнзимА, ацетил-КоА), углекислый газ и НАД • Н2:

2) в матриксе митохондрий уксусная кислота вступает в сложный цикл биохимических превращений, который получил название цикл трикарбоновых кислот или цикл Кребса. Немецкий биохимик Ханс Кребс за исследования аэробного обмена совместно с биохимиком Ф. Липманом был удостоен в 1953 г. Нобелевской премии.

Рис. 4.15. Процессы, происходящие в митохондриях:
А — энергетика митохондрии; Б — организация электронно-транспортной цепи окислительного фосфорилирования на мембранах митохондрий (клеточное дыхание): 1 — наружная мембрана; 2 — внутренняя мембрана; 3 — межмембранное пространство; 4 — матрикс митохондрий FqhFj — компоненты АТФ-синтетазного комплекса на внутренней мембране
Цикл Кребса состоит из восьми последовательных реакций, образующих круг (рис. 4.16). При окислении одной молекулы глюкозы происходит два оборота цикла Кребса.

Рис. 4.16. Схема цикла лимонной кислоты (цикл Кребса) 1
Итоговое уравнение одного оборота цикла Кребса следующее:

В результате сложных превращений из одной молекулы АцКо-А образуется три молекулы НАД • Н2, одна молекула ФАД • Н2 (еще один восстановленный переносчик атомов водорода — флавинадениндинукле- отид), одна молекула АТФ и две молекулы С02;
3) молекулы НАД • Н2 и ФАД -Н2 взаимодействуя с белками внутренней мембраны митохондрий, теряют атомы водорода, которые превращаются в ионы водорода Н + и электроны. Молекула НАД • Н2 окисляется при этом в НАД • Н+. С помощью специальной жирорастворимой витаминоподобной молекулы убихинона (коэнзим-QlO) и группы железосодержащих белков (цитохромы а, Ь, с), электроны переносятся по электронно-транспортной цепи во внутренней мембране митохондрии. Важнейшим ферментом, катализирующим перенос электронов по дыхательной цепи, является — цитохромоксидаза. В процессе этого переноса электроны теряют энергию, которая идет на перекачку ионов
Биология клетки / под ред.А. Ф. Никитина. 2-е изд. СПб. : Спецлит, 2015.
водорода через внутреннюю мембрану митохондрии в межмембранное пространство;
Механизмы образования АТФ в митохондриях
Все живые клетки обладают способностью превращать энергию окружающей среды в общий интермедиат — АТФ.
Клетка получает энергию за счет пищи, которую она усваивает из окружающей среды. Затем эта энергия должна быть превращена в форму, которую можно распределять по клетке. Общее решение этого вопроса (не только для митохондрий, но и для прокариотических клеток, также превращающих энергию) состоит в запасании энергии в форме универсального вещества, которое можно использовать в любой момент и в любом месте клетки.
Различные типы клеток отличаются деталями энергетической системы, однако общим свойством, характерным для всех клеток, является их способность превращать энергию окружающей среды в АТФ, который представляет собой общую молекулу, при необходимости обеспечивающую энергией отдельные химические реакции.
АТФ образуется двумя путями: в цитозоле и в митохондриях. Первый путь реализуется в цитозоле эукариотических клеток и у бактерий. При этом в результате гликолиза происходит распад глюкозы с образованием пирувата и образуются две молекулы АТФ. Эта реакция может происходить в анаэробных условиях (в отсутствие кислорода).
Второй путь является основным источником создания энергетических запасов и осуществляется в митохондриях эукариотических клеток. Процесс, при котором в митохондриях образуется АТФ, называется окислительным фосфорилированием и осуществляется с участием электрон-транспортной цепи. Пируват, высвобождающийся при гликолизе, поступает в матрикс (люмен) митохондрии, где он распадается и, реагируя с коэнзимом А, образует ацетил-КоА.
После этого, при участии цикла лимонной кислоты с освобождением атомов водорода, ацетильный остаток ацетил-КоА распадается, образуя двуокись углерода. Восстановление переносчика НАД+ в НАДН происходит при участии атомов водорода, и затем при окислении НАДН высвобождаются протон и электрон. Из матрикса ионы водорода (протоны) переносятся через мембрану в межмембранное пространство, а электроны перемещаются вдоль мембраны с участием нескольких транспортных белков. В результате по обе стороны мембраны создается градиент концентрации протонов.
Под действием этого градиента протоны переносятся через мембрану в обратном направлении. Это осуществляется при участии большого белкового комплекса АТФ-синтазы, которая и образует АТФ из АДФ и неорганического фосфата. Этот процесс носит название хемиосмос.
Митохондрии часто называют энергетическими центрами (эукариотических) клеток; они снабжают клетку энергией, необходимой для осуществления метаболических процессов и структурных изменений. Точнее говоря, они превращают энергию, поступающую из внешней среды в формы, которые могут быть использованы клеткой. Постоянство структуры и функции митохондрий у всех клеток эукариот убеждает в том, что эндосимбиоз, в результате которого они возникли, должен был произойти на самом начальном этапе эволюции эукариот.
АТФ образуется при хемиосмосе с участием системы протонного транспорта.
Видео урок цикл Кребса
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: